
向水性是植物根尖感知环境中的水势梯度并向着水势较高区域弯曲生长的生物学特性。这种特性对于植物有效地从土壤中吸收养分和水分,以及调整根系结构至关重要。Ca2+是植物生长发育和逆境响应的核心调控因子,一直是植物学研究领域的热点和挑战之一。虽然钙信号在植物向水性响应中起着重要的作用,但其确切的调控机制仍很不清楚。
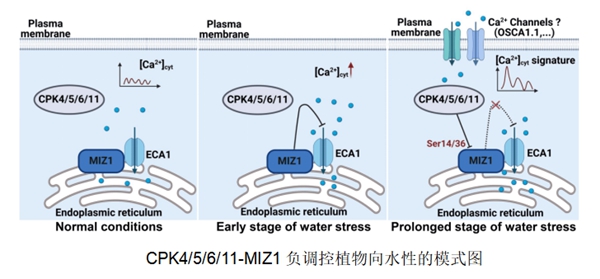
近日,生命科学学院王存教授团队在钙信号负调控植物向水性方面取得新进展。相关研究以“Arabidopsis CALCIUM-DEPENDENT PROTEIN KINASE4/5/6/11 negatively regulate hydrotropism via phosphorylation of MIZU-KUSSEI1”为题发表在《The Plant Cell》上。该研究利用拟南芥内质网定位的向水性特异蛋白MIZ1作为诱饵,对钙依赖性蛋白激酶CPK家族34个成员进行双分子荧光互补(BiFC)互作筛选发现,CPK4/5/6/11与MIZ1存在较强的相互作用,此外利用膜系统酵母双杂交、荧光素酶互补等多种互作手段验证了CPK4/5/6/11与MIZ1互作的真实性。进一步在水分胁迫培养基上进行表型实验发现,cpk4/5/6/11突变体对水势差的敏感性增加,根尖弯曲度增大。而过表达CPK5/11的转基因株系呈现出与miz1突变体相似的表型,对水势差敏感性显著降低,根尖基本不弯曲。
CPK作为蛋白激酶主要通过磷酸化下游底物实现信号的传递。通过体内/体外磷酸化实验发现,CPK5/11能够磷酸化修饰MIZ1,并且第14和36位的丝氨酸是其关键磷酸化位点。进一步通过高钙敏感缺陷酵母转运活性分析、转基因株系亚细胞定位和Western-blot等实验发现CPK介导的MIZ1的磷酸化不影响MIZ1的亚细胞定位和蛋白稳定性,而影响其对钙泵ECA1的转运活性的调控。为了进一步探究关键磷酸化位点的生物学功能,进行了体内钙信号变化测定和MIZ1点突变形式回补转基因株系的表型分析等实验,也表明了CPK介导的MIZ1Ser14/36磷酸化修饰对向水性中Ca2+信号转导至关重要。
综上所述,在正常情况下,[Ca2+]cyt保持相对静息水平。然而,在水分布不均匀的条件下,MIZ1与内质网Ca2+泵ECA1结合,抑制其转运活性,导致[Ca2+]cyt增加产生向水性所需的Ca2+信号,植物向水势较高的区域弯曲生长。在水分胁迫后期,CPK4/5/6/11感知细胞质Ca2+信号变化,磷酸化MIZ1以解除其对ECA1的抑制作用,进而导致细胞质钙流入内质网,[Ca2+]cyt水平降低,最终负向调控植物的向水性。研究结果揭示了Ca2+信号通过CPK4/5/6/11-MIZ1负向调控植物向水性的分子机制,为提高植物水分利用效率和促进农业可持续发展提供了理论依据。
博士后琚传凤为论文第一作者,王存教授为论文通讯作者。该研究得到了国家自然科学基金、中国博士后科学基金和中央高校基本科研业务费专项资金等项目的资助,作物抗逆与高效生产全国重点实验室技术平台以及生命科学学院技术平台提供了技术支持。

西北农林科技大学

西农深圳研究院